 PDF(2053 KB)
PDF(2053 KB)

十二烷酰肉碱和肉豆蔻酸对小鼠肺泡上皮细胞系MLE-12细胞功能的影响
马圆, 张婷, 姜志龙, 高佳萌, 钱宇豪, 陈智鸿
复旦学报(医学版) ›› 2025, Vol. 52 ›› Issue (03) : 333-342.
PDF(2053 KB)

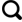 PDF(2053 KB)
PDF(2053 KB)
十二烷酰肉碱和肉豆蔻酸对小鼠肺泡上皮细胞系MLE-12细胞功能的影响
Effect of dodecanoylcarnitine and myristoleic acid on the cellular function of mouse alveolar epithelial cell line of MLE-12
目的: 探究十二烷酰肉碱(dodecanoylcarnitine,DA)和肉豆蔻酸(myristoleic acid,MA)对小鼠肺泡上皮系MLE-12细胞功能的影响及其作用机制。方法: 通过IL-4刺激MLE-12细胞构建炎症模型,采用ELISA检测细胞上清液中DA、MA以及鞘氨醇-1-磷酸(sphingosine-1-phosphate,S1P)表达水平;用DA和MA分别干预MLE-12细胞,RT-PCR、流式细胞术检测炎症因子IL-6和肿瘤坏死因子α(tumor necrosis factor-α,TNF-α)表达变化及细胞内活性氧(reactive oxygen species,ROS)水平,进一步通过Western blot检测p-38 MAPK蛋白激酶(p38 mitogen activated protein kinase,p-38 MAPK)和src同源结构域2的酪氨酸磷酸酶1(src homology 2 domain containing phosphatase 1,SHP-1)等关键蛋白表达情况;使用S1PR2的抑制剂JTE-013预处理MLE-12细胞后重复上述实验,系统评估S1PR2信号通路在DA/MA介导的炎症反应及氧化应激调控中的分子机制。结果: IL-4刺激显著上调MLE-12细胞中DA、MA及S1P水平(P<0.05)。DA/MA干预组炎症因子IL-6、TNF-α表达量较对照组明显升高(P<0.05),同时ROS水平较对照组也有所增加(P<0.05)。Western blot结果显示DA/MA促进SHP-1去磷酸化及p-p38 MAPK活化,而JTE-013预处理可完全逆转上述效应(P<0.05)。结论: 哮喘相关代谢物DA和MA通过激活S1PR2受体,促进SHP-1去磷酸化和p-p38 MAPK通路活化,加剧MLE-12细胞的炎症及氧化应激反应,本研究揭示了S1PR2在该通路中的核心调控作用。
Objective: To explore the effects of dodecanoylcarnitine (DA) and myristoleic acid (MA) on the function of mouse alveolar epithelial cell line MLE-12 and their underlying mechanisms. Methods: An inflammatory model was established by stimulating MLE-12 cells with IL-4. The expression levels of DA, MA, and sphingosine-1-phosphate (S1P) in the cell supernatant were detected by ELISA. MLE-12 cells were separately intervened with DA and MA. RT-PCR and flow cytometry were used to detect the expression changes of inflammatory factors IL-6 and tumor necrosis factor-α (TNF-α) and the level of intracellular reactive oxygen species (ROS). Additionally, Western blot was performed to detect the expression of key proteins such as p38 mitogen-activated protein kinase (p-38 MAPK) and src homology 2 domain-containing phosphatase 1 (SHP-1). To explore the role of S1PR2 in the effects of DA and MA, MLE-12 cells were pretreated with the S1PR2 inhibitor JTE-013, and the above experiments were repeated. Results: IL-4 stimulation significantly upregulated the levels of DA, MA, and S1P in MLE-12 cells (P<0.05). DA/MA treatment groups exhibited significantly increased expression of IL-6 and TNF-α compared with the control group (P<0.05), along with elevated ROS levels (P<0.05). Western blot analysis revealed that DA/MA promoted SHP-1 dephosphorylation and phosphorylated p38 MAPK activation in MLE-12 cells. Notably, JTE-013 pre-treatment completely reversed these effects (P<0.05). Conclusion: Asthma-related metabolites DA and MA exacerbate the inflammatory and oxidative stress responses of MLE-12 cells by activating the S1PR2 receptor, promoting the dephosphorylation of SHP-1 and the activation of the p-p38 MAPK pathway. This study reveals the core regulatory role of S1PR2 in this pathway as well.
哮喘 / 代谢 / 十二烷酰肉碱(DA) / 肉豆蔻酸(MA) / JTE-013 / 小鼠 {{custom_keyword}} /
asthma / metabolism / dodecanoylcarnitine (DA) / myristoleic acid (MA) / JTE-013 / mouse {{custom_keyword}} /
表 1 qRT-PCR引物序列Tab 1 qRT-PCR primers for analysis |
| Gene | Primers(5’-3’) |
| IL-6 | Forward:CACAAGTCCGGGAGGAGAC |
| Reverse:TCCACGATTTCCCAGAGAAC | |
| TNF-α | Forward:CCAAATGGCCTCCCTCTCAT |
| Reverse:GGTGGTTTGCTACGACGTGG | |
| S1PR2 | Forward:CATCTTACTGGCTATCGTGGCTCTG |
| Reverse:GCGTCTGAGGACCAGCAACATC | |
| GAPDH | Forward:GGCAAATTCCATGGCACCGT |
| Reverse:GCATCGCCCCACTTGATTTT |
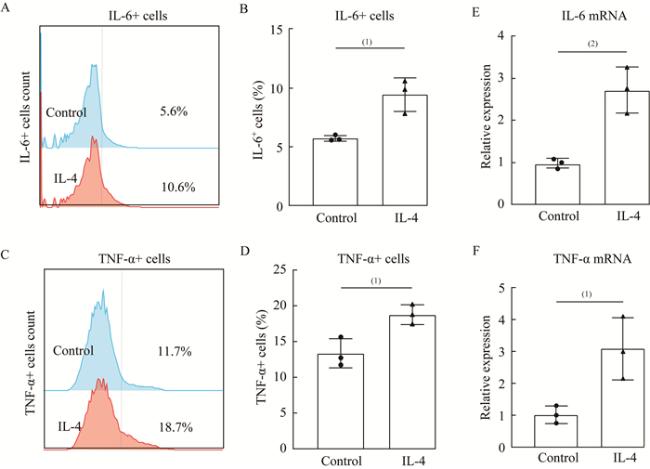
图 1 构建IL-4诱导的MLE-12细胞炎症模型Fig 1 Generation of of IL-4-induced MLE-12 cell inflammation model A and B: The expression of IL-6 in MLE-12 cells between IL-4 and control group was measured by flow cytometry and quantified; C and D: The expression of TNF-α in MLE-12 cells between IL-4 and control group was measured by flow cytometry and quantified; E and F: IL-6 and TNF-α mRNA expression was detected by qRT-PCR. n=3 per group, (1)P<0.05, (2)P<0.01 by two-tailed Student t-test, |
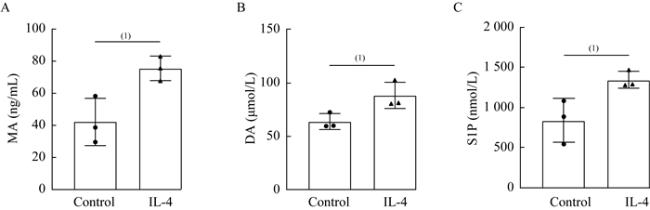
图 2 IL-4诱导MLE-12分泌DA、MA和S1PFig 2 The secretion of DA, MA and S1P of MILE-12 cells induced by IL-4 A: The value of MA in the supernatant of MLE-12 cells was detected by ELISA; B: The value of DA in the supernatant of MLE-12 cells was detected by ELISA; C: The value of S1P in the supernatant of MLE-12 cells was detected by ELISA. MA: Myristoleic acid; DA: Dodecanoylcarnitine; S1P: Sphingosine-1-phosphate. n=3 per group, (1)P<0.05 by two-tailed Student t-test, |
图 3 JTE-013有效抑制DA和MA诱导的MLE-12细胞S1PR2表达Fig 3 JTE-013 effectively inhibited the expression of S1PR2 in MLE-12 cells induced by DA and MA A and B: The expression of S1PR2 in MLE-12 cells among control, DA and DA+JTE-013 group was measured by flow cytometry and quantified; C and D: The expression of S1PR2 in MLE-12 cells among control, MA and MA+JTE-013 group was measured by flow cytometry and quantified; E and F: S1PR2 mRNA expression was detected by qRT-PCR. n=3 per group, (1)P<0.05, (2)P<0.01, (3)P<0.001, (4)P<0.000 1 by two-tailed Student t-test, |
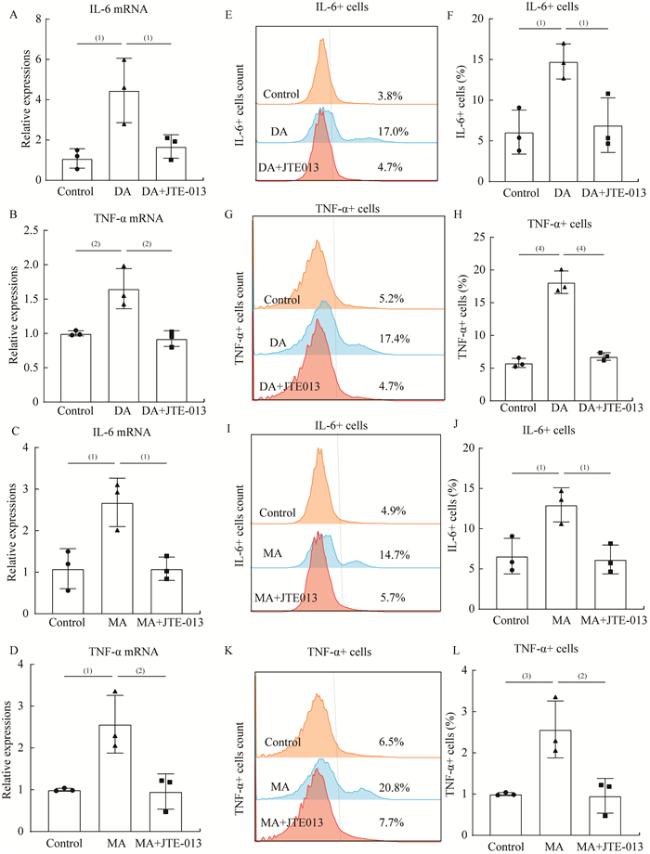
图 4 JTE-013有效抑制DA和MA促进的MLE-12细胞的炎症分泌Fig 4 JTE-013 inhibited DA-induced and MA-induced allergic inflammatory responses in MLE-12 A-D: IL-6 and TNF-α mRNA expression was detected by qRT-PCR; E-H: The expression of IL-6 and TNF-α in MLE-12 cells among control, DA and DA+JTE-013 group was measured by flow cytometry and quantified; I-L: The expression of IL-6 and TNF-α in MLE-12 cells among control, MA and MA+JTE-013 group was measured by flow cytometry and quantified. n=3 per group, (1)P<0.05, (2)P<0.01, (3)P<0.001, (4)P<0.000 1 by two-tailed Student t-test, |
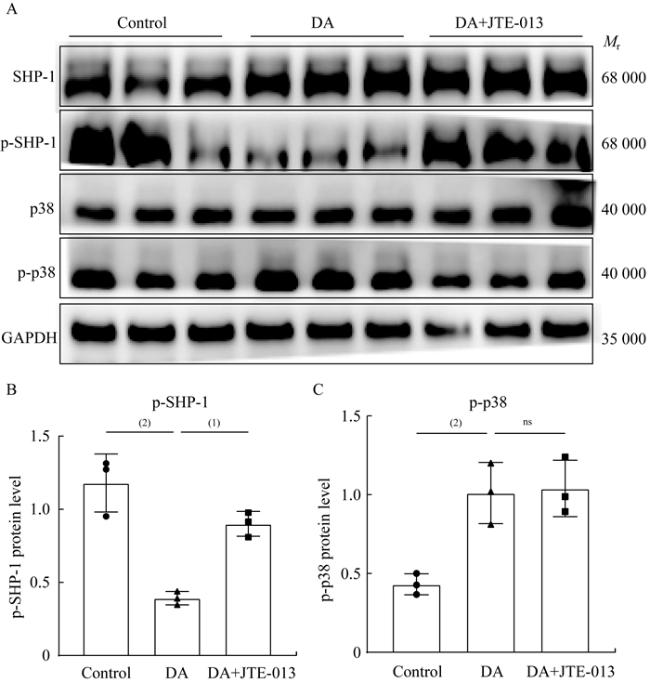
图 6 DA通过S1PR2/SHP-1/p38 MAPK信号通路介导炎症反应激活Fig 6 DA-mediated inflammatory activation was dependent on the S1PR2/SHP-1/p38 MAPK signaling pathway A: Protein expression level among different groups in MLE-12 was detected by Western blot; B and C: Quantitative analysis of p-SHP-1 and p-p38 protein expression in MLE-12. n=3 per group, (1) P<0.01, (2)P<0.001 by two-tailed Student t-test, |
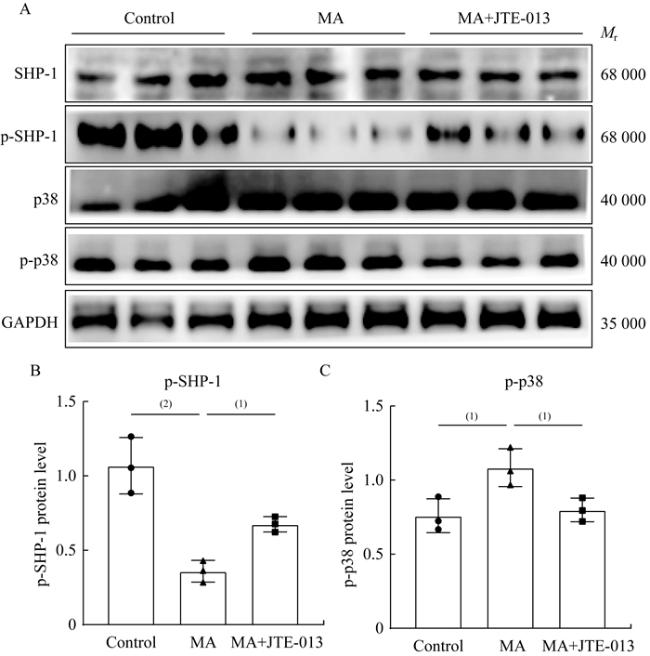
图 7 MA通过S1PR2/SHP-1/p38 MAPK信号通路介导炎症反应激活Fig 7 MA-mediated inflammatory activation was dependent on the S1PR2/SHP-1/p38 MAPK signaling pathway A: Protein expression level among different groups in MLE-12 was detected by Western blot. B and C: Quantitative analysis of p-SHP-1 and p-p38 protein expression in MLE-12. n=3 per group, (1)P<0.05, (2) P<0.001 by two-tailed Student t-test, |
| 1 |
PAPI A, BRIGHTLING C, PEDERSEN SE, et al. Asthma[J]. Lancet, 2018, 391(10122): 783- 800.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 2 |
WANG G, MCDONALD VM. Contemporary concise review 2020: asthma[J]. Respirology, 2021, 26(8): 804- 811.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 3 |
Global burden of chronic respiratory diseases and risk factors, 1990-2019: an update from the Global Burden of Disease Study 2019[J]. EClinicalMedicine, 2023, 59: 101936.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 4 |
周新, 张旻. 中国支气管哮喘防治指南(2020年版)解读[J]. 诊断学理论与实践, 2021, 20(2): 138- 143.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 5 |
CRESTANI E, HARB H, CHARBONNIER LM, et al. Untargeted metabolomic profiling identifies disease-specific signatures in food allergy and asthma[J]. J Allergy Clin Immunol, 2020, 145(3): 897- 906.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 6 |
ZHU T, MA Y, WANG J, et al. Serum metabolomics reveals metabolomic profile and potential biomarkers in asthma[J]. Allergy Asthma Immunol Res, 2024, 16(3): 235- 252.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 7 |
IGUCHI K, OKUMURA N, USUI S, et al. Myristoleic acid, a cytotoxic component in the extract from Serenoa repens, induces apoptosis and necrosis in human prostatic LNCaP cells[J]. Prostate, 2001, 47(1): 59- 65.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 8 |
REINKE SN, NAZ S, CHALECKIS R, et al. Urinary metabotype of severe asthma evidences decreased carnitine metabolism independent of oral corticosteroid treatment in the U-BIOPRED study[J]. Eur Respir J, 2022, 59(6): 2101733.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 9 |
LOCHNER M, BEROD L, SPARWASSER T. Fatty acid metabolism in the regulation of T cell function[J]. Trends Immunol, 2015, 36(2): 81- 91.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 10 |
CHIURCHIÙ V, LEUTI A, MACCARRONE M. Bioactive lipids and chronic inflammation: managing the fire within[J]. Front Immunol, 2018, 9(1): 38.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 11 |
PARK SJ, IM DS. Blockage of sphingosine-1-phosphate receptor 2 (S1P 2) attenuates allergic asthma in mice[J]. Brit J Pharmacol, 2019, 176(7): 938- 949.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 12 |
蒋艳, 张韶泽, 曾丹, 等. 1-磷酸鞘氨醇在呼吸系统疾病中的研究进展[J]. 中国医师杂志, 2021, 23(12): 5.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 13 |
BROUWERS H, VON HEGEDUS J, TOES R, et al. Lipid mediators of inflammation in rheumatoid arthritis and osteoarthritis[J]. Best Pract Res Clin Rheumatol, 2015, 29(6): 741- 755.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 14 |
FROMMER KW, SCHÄFFLER A, REHART S, et al. Free fatty acids: potential proinflammatory mediators in rheumatic diseases[J]. Ann Rheum Dis 2015, 74(1): 303-310.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 15 |
FROMMER KW, HASSELI R, SCHÄFFLER A, et al. Free fatty acids in bone pathophysiology of rheumatic diseases[J]. Front Immunol, 2019, 10: 2757.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 16 |
JIANG J, WOLK A, VESSBY B. Relation between the intake of milk fat and the occurrence of conjugated linoleic acid in human adipose tissue[J]. Am J Clin Nutr, 1999, 70(1): 21- 27.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 17 |
COUVREUR S, HURTAUD C, LOPEZ C, et al. The linear relationship between the proportion of fresh grass in the cow diet, milk fatty acid composition, and butter properties[J]. J Dairy Sci, 2006, 89(6): 1956- 1969.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 18 |
REINKE SN, NAZ S, CHALECKIS R, et al. Urinary metabotype of severe asthma evidences decreased carnitine metabolism independent of oral corticosteroid treatment in the U-BIOPRED study[J]. Eur Respir J, 2022, 59(6): 2101733.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 19 |
IGUCHI K, OKUMURA N, USUI S, et al. Myristoleic acid, a cytotoxic component in the extract from Serenoa repens, induces apoptosis and necrosis in human prostatic LNCaP cells[J]. Prostate, 2001, 47(1): 59- 65.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 20 |
KWON JO, JIN WJ, KIM B, et al. Myristoleic acid inhibits osteoclast formation and bone resorption by suppressing the RANKL activation of Src and Pyk2[J]. Eur J Pharmacol, 2015, 768: 189- 198.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 21 |
QUAN LH, ZHANG C, DONG M, et al. Myristoleic acid produced by enterococci reduces obesity through brown adipose tissue activation[J]. Gut, 2020, 69(7): 1239- 1247.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 22 |
FUJIOGI M, CAMARGO CA, JR, RAITA Y, et al. Integrated associations of nasopharyngeal and serum metabolome with bronchiolitis severity and asthma: a multicenter prospective cohort study[J]. Pediatr Allergy Immunol, 2021, 32(5): 905- 916.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 23 |
ZHANG J, SOMANI AK, SIMINOVITCH KA. Roles of the SHP-1 tyrosine phosphatase in the negative regulation of cell signalling[J]. Semin Immunol, 2000, 12(4): 361- 378.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 24 |
LIPAO, BADOUR K, SIMINOVITCH KA, et al. Nonreceptor protein-tyrosine phosphatases in immune cell signaling[J]. Annu Rev Immunol, 2007, 25(1): 473- 523.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 25 |
KAMATA T, YAMASHITA M, KIMURA M, et al. src homology 2 domain-containing tyrosine phosphatase SHP-1 controls the development of allergic airway inflammation[J]. J Clin Invest, 2003, 111(1): 109- 119.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 26 |
JANG MK, KIM SH, LEE KY, et al. The tyrosine phosphatase, SHP-1, is involved in bronchial mucin production during oxidative stress[J]. Biochem Biophys Res Commun, 2010, 393(1): 137- 143.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| {{custom_ref.label}} |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
作者贡献声明 马圆 实验实施,论文撰写和修订。张婷 实验设计,数据分析,论文撰写。姜志龙 实验方法设计。高佳萌,钱宇豪 实验数据分析。陈智鸿 实验方法设计,论文指导和修订。
利益冲突声明 所有作者均声明不存在利益冲突。
PDF(2053 KB)
 表 1 qRT-PCR引物序列
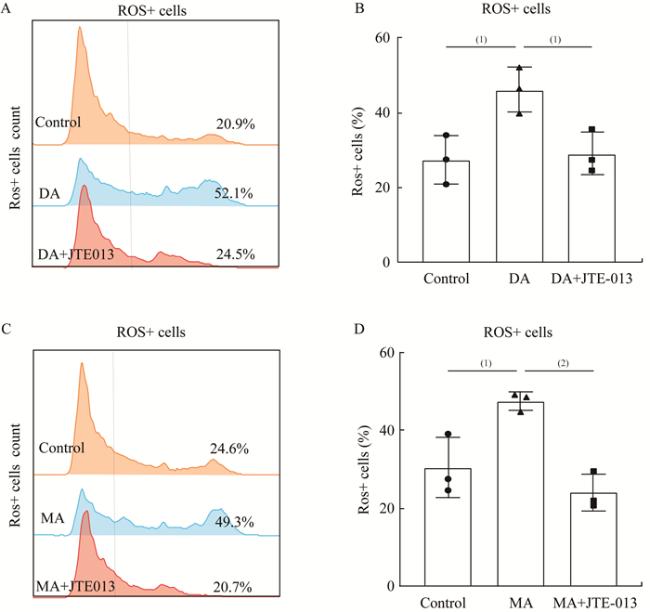
表 1 qRT-PCR引物序列 图 1 构建IL-4诱导的MLE-12细胞炎症模型图 2 IL-4诱导MLE-12分泌DA、MA和S1P图 3 JTE-013有效抑制DA和MA诱导的MLE-12细胞S1PR2表达图 4 JTE-013有效抑制DA和MA促进的MLE-12细胞的炎症分泌图 5 JTE-013抑制DA和MA促进的MLE-12组胞氧化应激图 6 DA通过S1PR2/SHP-1/p38 MAPK信号通路介导炎症反应激活图 7 MA通过S1PR2/SHP-1/p38 MAPK信号通路介导炎症反应激活
图 1 构建IL-4诱导的MLE-12细胞炎症模型图 2 IL-4诱导MLE-12分泌DA、MA和S1P图 3 JTE-013有效抑制DA和MA诱导的MLE-12细胞S1PR2表达图 4 JTE-013有效抑制DA和MA促进的MLE-12细胞的炎症分泌图 5 JTE-013抑制DA和MA促进的MLE-12组胞氧化应激图 6 DA通过S1PR2/SHP-1/p38 MAPK信号通路介导炎症反应激活图 7 MA通过S1PR2/SHP-1/p38 MAPK信号通路介导炎症反应激活/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}